FIGURE 12.29 A succession of plant species on abandoned fields in North Carolina. Pioneer species consist of a variety of annual plants. This successional stage is followed by communities of perennials and grasses, shrubs, softwood trees and shrubs, and finally, hardwood trees and shrubs. This succession takes about 120 years from the pioneer stage to the climax community. Image Copyright: Michael Pidwirny.
FIGURE 12.34 Yellow Mountain-Avens (Dryas drummondii) is a pioneer species associated with plant succession on glacial deposits. This nitrogen-fixing plant species plays a facilitative role in this succession by enriching the soil with nitrogen. This enrichment allows subsequent colonization by Cottonwood (Populus trichocarpa) and Sitka Spruce (Picea sitchensis) trees. Image Source: Wikimedia Commons, photo by Kurt Stueber. This image is licensed under the Creative Commons Attribution-Share Alike 3.0 Unported license.
The term plant succession describes a directional, non-seasonal, cumulative change in the types of plant species that occupy a given area through time. This process occurs on a timescale longer than a single growing season. Succession involves the processes of colonization, establishment, and extinction that act on the participating plant species. Most successions contain several stages, each characterized by the species that dominate at that point in time. Most successions begin when an area is partially or completely cleared of vegetation due to a disturbance. Shortly after the disturbance, the first wave of colonizing plants, or the pioneer community, establishes itself (Figure 12.28). The pioneer community usually consists of short-lived, small, fast-growing plant species. Successive stages of plant re-growth then follow this event on the disturbed site. Through this temporal sequence, there is a general trend of the species involved having longer lifespans, larger physical sizes, and slower growth rates. From a hypothetical standpoint, succession stops when species composition no longer changes with time. This stage is known as the climax community.
The concept of a climax community assumes that the plants colonizing and establishing themselves in a given region can achieve a stable equilibrium over time. The idea that succession culminates in a climax community has a long history in the fields of biogeography and ecology. One of the earliest proponents of this theory was Frederic Clements, who studied succession at the beginning of the 20th century. Clements believed that most plant communities undergoing succession reach the climax state. In the 1920s, scientists began questioning the notion of a climax state because of mounting field evidence. By 1950, many scientists began to see succession as a process that rarely attains a stable equilibrium or climax community. The reason equilibrium is rarely reached is related to the nature of the disturbance. Clements’s succession theory was based on the assumption that plant communities were seldom disturbed. Yet numerous studies of natural vegetation conducted after he published his idea indicated that disturbance was a frequent phenomenon in most plant communities. Because of its high frequency, most plant communities rarely reach a stage in their development where species composition reaches a steady-state climax community.
Studies of succession also revealed that disturbance acts on plant communities at a variety of spatial and temporal scales. Further, the effect of disturbance is not always 100%. Most disturbance events remove only a part of the previous plant community. As a result of these new ideas, plant communities are now generally seen as composed of numerous patches of various sizes at different levels of successional development. A particular patch in a larger community may undergo repeated disturbance events that do not allow it to continue past the pioneer stage. However, the general characteristic of the larger plant community around the patch may be late-successional.
Abandoned Field to Oak Forest
Dwight Billings did one of the earliest studies of plant succession in the 1930s. In this investigation, Billings examined the succession of plant species on abandoned agricultural fields in North Carolina. Billings studied several fields that had been deserted for just a few years to as long as about 150 years. From observations of the plant communities in these sites, Billings could construct a detailed successional sequence.
The first stage of succession was characterized by the colonization of bare ground and nutrient-poor soils by annual plant species (Figure 27.29). These annual species have short life spans (one growing season), rapid maturity, and produce numerous small, easily dispersed seeds. The annuals were then quickly replaced by biennial plants and grasses the following year. The annual, biennial, and grass types of plants are often called pioneer species in plant succession. After about 3 to 4 years, the biennial and grass species gave way to perennial herbs and shrubs. These plants live for many years and can reproduce several times during their lifetimes.
After about 5 to 15 years, the sites were colonized by several softwood tree species, including Loblolly Pine (Pinus taeda), Shortleaf Pine (Pinus echinata), Virginia Pine (Pinus virginiana), and Sweetgum (Liquidambar spp.). As the softwoods increased in numbers and grew in height, they began forming a forest canopy. This canopy reduces the amount of light reaching the forest floor. The resulting shaded understory conditions caused the exclusion of many light-loving perennial herb and shrub species. Low light conditions also inhibited the germination of pine seedlings. Perennial herb and shrub species adapted to low light conditions were now taking over the ground cover. The canopy also altered the microclimate near ground level in the habitat. It was now more humid, with milder temperatures and less wind. These conditions, along with the development of a soil litter layer, allowed the germination of hardwood species, such as Oak (Quercus spp.) and various Hickory (Carya spp.) species. The seedlings of these tree species can also tolerate low light levels.
By about 50 to 75 years after the initial colonization of the pioneer species, the hardwoods began to replace the softwoods in the developing forest. At this stage in the succession, the pines had maximum heights of about 25 meters (75 feet), while the oaks and hickories were, on average, about 10 meters (30 feet) tall. Because of their shorter life spans (50 years), many softwood species began dying out. Subdominant hardwood trees would fill the gaps produced by the softwood species' deaths. Hardwood species, like Oak and Hickory, can live for over 100 years. Sites more than 100 years old were found to be dominated by mature Oak forests (Figure 27.30).
Types of Plant Successions
At the most basic level, we can classify plant successions into two types: primary succession and secondary succession. Primary succession occurs when plants establish on land that was not previously vegetated or in habitats where disturbance has killed all the seeds stored in the soil surface layers (Figure 27.31). Habitats undergoing primary succession include vegetation that develops on volcanic deposits, rock outcrops, sand dunes, newly created river sandbars, newly formed oceanic islands, and exposed ground surfaces de-vegetated by glaciers. Secondary succession involves the invasion of a site by plants on the ground that was previously vegetated (Figure 27.32). The established plants often come from seeds stored in the site’s soil. Removal of past vegetation may be caused by natural or human-mediated disturbances such as fire, logging, cultivation, or weather disturbances like strong winds, tornadoes, and hurricanes.
Successions can also be defined by the factors driving the changes in species composition. An allogenic succession is caused by a change in environmental conditions that, in turn, influences the composition of the plant community. In Cornwall, England, observations on the Fal River estuary suggest that silt deposition may be causing an allogenic succession from salt marsh to woodland (Ranwell, 1974). Measurements indicate sedimentation rates of about 1 cm per year on the mud flats that are found 15 kilometers (9 miles) into the estuary. Over the last 100 years, this salt marsh has increased its elevation and extended seaward by 800 meters (2600 feet). The adjacent woodland has advanced into the salt marsh, reaching its landward limit.
Succession can also be driven by the species found in the plant community. In this type of succession, the plant community alters the abiotic environment through its presence. This environmental modification then facilitates a shift in species composition. This type of succession can be called an autogenic succession. In the coniferous forests of coastal Oregon, Washington, and British Columbia, Red Alder (Alnus rubra) is a fast-growing and short-lived early colonizer of disturbed sites where vegetation has been removed. Red Alder can also form symbiotic relationships with nitrogen-fixing bacteria. This association causes soil nitrogen levels to increase dramatically over time. With higher soil nitrogen levels, Douglas Fir (Pseudotsuga menziesii) seedlings become more successful in colonizing and establishing themselves on these sites. Eventually, the Douglas Fir becomes more dense and grows tall enough to cause the competitive demise of Red Alder by limiting light transmission to the forest floor (Figure 27.33).
Abiotic and Biotic Changes During Succession
Analyzing the research on numerous succession examples in various habitat types reveals some interesting patterns. These patterns are fascinating because they provide insights into the possible operating processes. In terms of biotic and abiotic factors, the following differences can be identified when comparing early with late successional stages:
- Pioneer species tend to be small, short-lived, and fast-growing.
- Late successional species are adapted to long lifespans, have large biomass, and grow slowly.
- The general morphology and physiology of late successional species tend to be more complex. They have many more adaptations required for survival.
- Late successional species, especially their seedlings and young plants, tend to have high photosynthetic efficiency at low light levels. This requirement requires a heavy investment in nitrogen to produce very efficient chloroplasts.
- The quantity of nitrogen available for uptake increases substantially when one moves from early to late stages in most successions.
- Early successional species tend to produce small seeds with dispersal mechanisms that carry them great distances. However, to be successful, these seeds must colonize sites free of competitors.
- Late successional species typically have large, poorly dispersed seeds. These seeds contain enough resources to produce very competitive seedlings upon germination, especially for light.
- Early successional species tend to be good competitors for soil nutrients and water but poor competitors for light.
- The decomposition of organic matter in the early stages tends to be rapid, with considerable loss from the community. In late stages, decomposition rates are slow, with most of the released nutrients returned to the plants.
- Site characteristics tend to be quite extreme in the early stages of succession compared to the late stages. The developing plant community moderates abiotic conditions.
- Organisms involved in the decomposition process become more abundant and more varied with time.
- Plant species diversity becomes greater over time since the start of the succession.
- Regarding plant adaptive strategies, early colonizers are ruderals, while late successional species belong to the competitive strategy.
Succession Mechanisms
In the late 1970s, ecologists J.H. Connell and R.O. Slatyer (1977) published an excellent survey of plant succession research. Considering their review of numerous past studies, Connell and Slatyer suggested that the processes involved in succession can be grouped into three general models: Facilitation, Tolerance, and Inhibition. They also suggested that most successions seem to involve processes from more than one model.
Facilitation Model of Succession
The defining feature of the facilitation model of succession is that the developing plant community changes the abiotic environment. These changes make the environment more favorable for plant species that were previously unable to colonize the site. Once colonized, the new species displaces the early site-modifying species through resource competition.
A great example of this form of succession was documented at Glacier Bay, Alaska. In 1916, William Cooper established long-term study plots in recently deglaciated areas. At this site, lichens, cyanobacteria, fungi, and mosses were the first organisms to colonize the bare ground and glacial till. Together, these organisms form thin mats that cover the ground. Under these mats, the glacial deposits begin to develop into proper soils due to the accumulation of organic matter, increased soil moisture retention, and reduced soil erosion. By about 20 to 50 years, the developing soils support a variety of grasses, herbaceous plants, and small shrubs on their surface. Two common colonizing shrubs at this stage in the succession are the Yellow Mountain-Avens (Dryas drummondii) and Mountain Alder (Alnus crispa) (Figure 27.34). Both of these plant species facilitate succession by altering the nutrient status of the developing soil. In both of these shrubs, nitrogen-fixing bacteria live symbiotically in root nodules. In exchange for photosynthetically produced carbohydrates, these bacteria provide the plants with nitrogen (nitrate). The presence of the bacteria also dramatically increases the quantity of growth-limiting nitrogen in the soil. This increase in soil nitrogen allows for the successful colonization of trees like Cottonwood (Populus trichocarpa) and Sitka Spruce (Picea sitchensis). These trees can only establish themselves on sites with sufficient quantities of soil nitrogen required for growth. The establishment of trees on the developing site also reduces the amount of light available on the ground surface. Consequently, this reduction in light causes the competitive exclusion of many grasses, herbaceous plants, and small shrubs growing beneath the tree canopy.
Tolerance Model of Succession
The tolerance model of succession suggests that a predictable sequence arises because different species employ different strategies to exploit resources. In this model, all colonizing plant species can survive as adults under the environmental conditions at the beginning of the succession. As succession progresses, soil and light resources become increasingly limited as the plant community develops and competition intensifies. Species that dominate in the early stages of succession tend to be small, easily dispersed, and have rapid growth. Over time, these species are replaced in dominance by larger, long-lived, slow-growing plants. These late succession species can tolerate lower resource levels due to competition and can grow to maturity in the presence of early species, eventually outcompeting them. This model is typical of the succession dynamics seen in some forested communities.
Inhibition Model of Succession
The inhibition model of succession also suggests that all species can survive as adults in the site during the early stages of the succession. It also indicates that the various species are unaffected by competitive interactions for soil nutrients or light. However, the species involved in the succession do compete for space. Further, these spaces only open up to a potential colonist when the present occupant is removed because of death. Death can occur from physical factors or biological interactions like disease or predation. In most successions, long-lived species become dominant with time because they gradually replace short-lived early colonists when they die.

FIGURE 12.28 Purple colored Fireweed (Epilobium angustifolium) is a common pioneer plant species that quickly colonizes temperate forest habitats that have been disturbed by fire or logging. Image Source: Wikimedia Commons, United States National Park Service, photo by Jim Peaco.
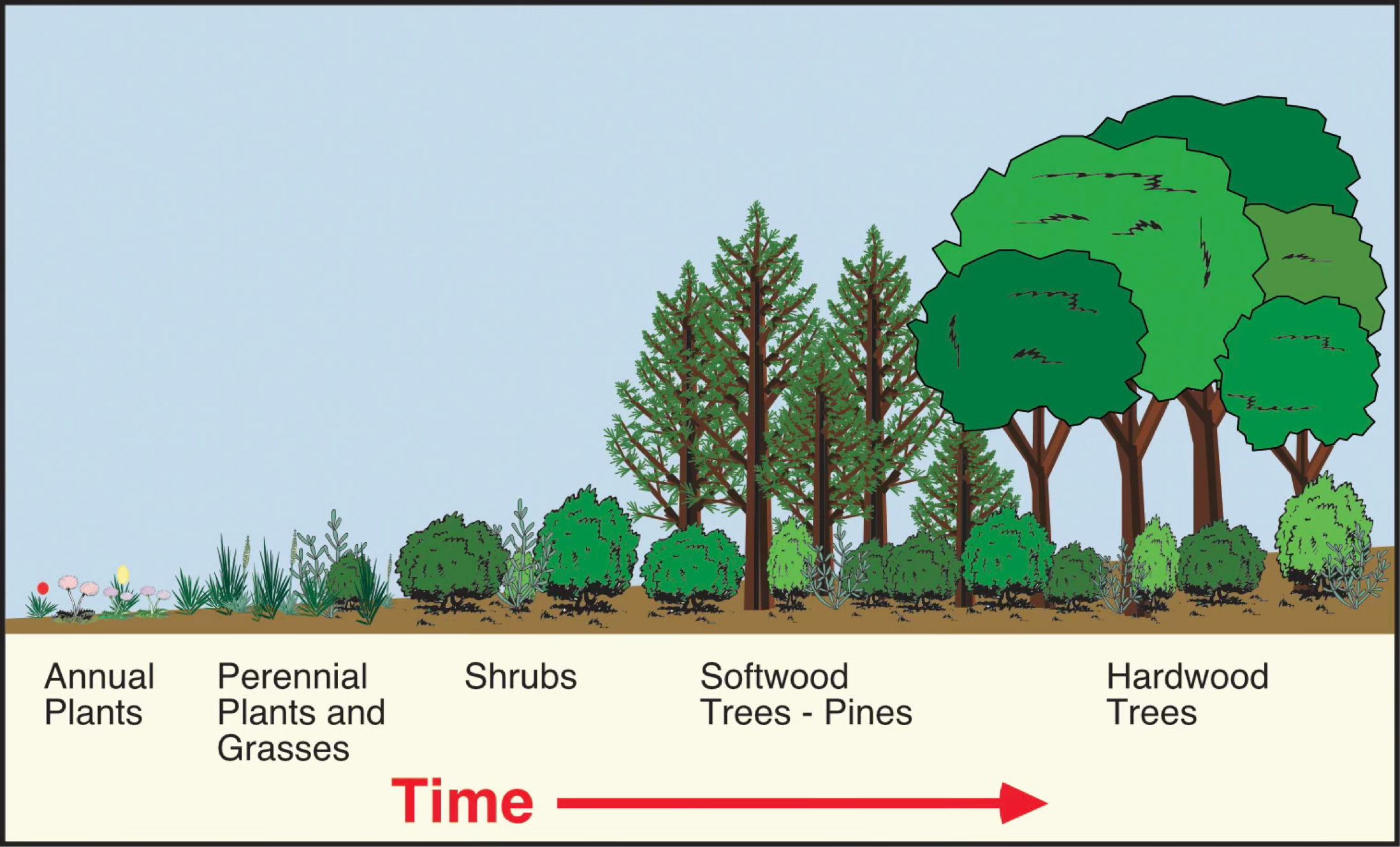

FIGURE 12.30 Very little light passes through the canopy of a mature oak-dominated forest. This environmental condition can cause the death of softwood species that grow best in intense sunlight. Image Source: Wikimedia Commons, photo by Brian Stansberry. This image is licensed under the Creative Commons Attribution-Share Alike 3.0 Unported license.


FIGURE 12.32 Plant life is returning to a forest site that was burned one year earlier. Some of the new plant life came from seeds stored in the soil that were not destroyed by the fire. Image Copyright: Michael Pidwirny.


FIGURE 12.31 The eruption of Mount Saint Helens on May 18, 1980, destroyed all life over a large region. Because this disturbance destroyed the soil seed bank of this area, this succession would be considered a primary succession. The image above shows that life has returned to the slopes of Mount Saint Helens. This photo was taken on July 31, 2007. Image Source: Wikimedia Commons, photo by Art Bromage. This image is licensed under the Creative Commons Attribution-Share Alike 2.0 Generic license.

FIGURE 12.33 Red Alder (Alnus rubra) is a tree that can symbiotically fix nitrogen with the help of certain species of actinomycetes. This relationship leads to an increase in soil nitrogen content over time. Shade-tolerant Douglas Fir (Pseudotsuga menziesii) seedlings are most successful in establishing themselves when the nitrogen concentration in the soil becomes quite plentiful. Once established, the Douglas Fir trees grow until they tower over and shade the shorter Red Alder. This shading reduces the light available for photosynthesis, eventually causing the Red Alder to die out. Image Source A: Wikimedia Commons, photo by Jeff Taylor. Image Copyright B: Michael Pidwirny.